
8. Țesutul muscular
Cuprins:
8.1. Introducere8.2. Mușchiul striat scheletic
8.3. Energia necesară contracției musculare
8.4. Funcțiile mușchiului neted și cardiac
8.1. Introducere
Țesutul muscular – unul dintre cele 4 țesuturi de bază, care se distinge prin capacitatea sa de contracție și posibilitatea de a efectua lucru mecanic.
Celula musculară (sau fibra musculară) are o formă alungită și reprezintă unitatea structurală a țesutului muscular. Acești termeni anatomici sunt similari, interschimbabili.
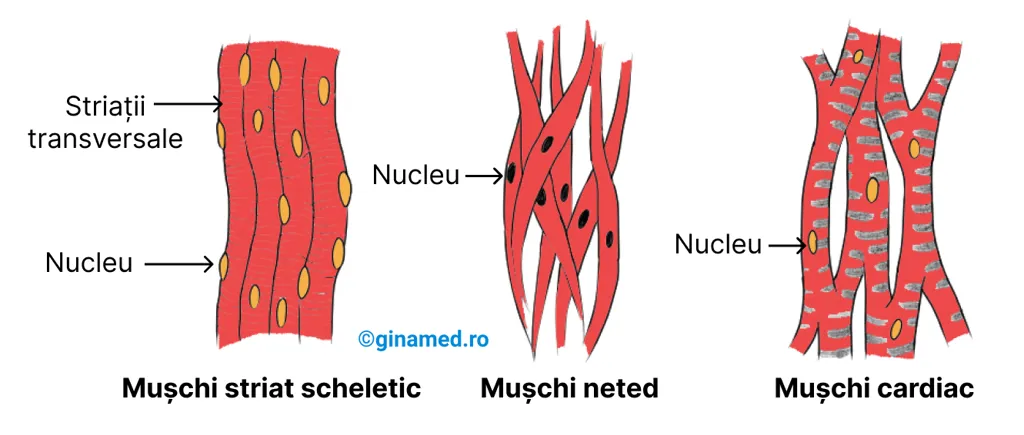
Există 3 tipuri de țesuturi musculare ale căror particularități (forme, funcții variate, etc.) sunt cuprinse în tabelul de mai jos:
| Caracteristică | Mușchi striat scheletic | Mușchi neted (nestriat) | Mușchi cardiac |
|---|---|---|---|
| Localizare | cel mai frecvent întâlnit tip de mușchi; inserat pe oase, atașat scheletului; unitatea mușchi-țesut osos facilitează mișcarea corpului | așezat în straturi în componența tubului digestiv, tractului respirator, peretele intestinelor, al uterului, al anumitor vase de sânge, în al anumitor canale (ducte), în viscere, etc. | în peretele inimii, contribuind la pomparea sângelui |
| Tip de control | voluntar | involuntar | involuntar |
| Forma fibrelor | alungite (foarte lungi), cilindrice, cu capete rotunjite; în realitate, fiecare celulă musculară este echivalentul a zeci sau sute de celule fuzionate și care sunt denumite tipic fibre musculare; un număr mare din aceste fibre alcătuiesc organul denumit „mușchi” | subțiri, alungite, fusiforme contractile, cu capete ascuțite | alungite, cilindrice, ramificate; proteinele contractile au o organizare mai complexă comparativ cu mușchiul neted; prezintă conexiuni speciale în peretele inimii prin joncțiuni „gap” la nivelul discurilor intercalare |
| Striații | prezente | absente | prezente |
| Număr de nuclei pe fibră | mulți | unul | unul |
| Poziția nucleilor în celulă | periferici | central | central |
| Viteza contracției | cel mai rapid | cel mai lent | intermediar |
| Capacitatea de a rămâne contractat | cea mai mică | cea mai mare | intermediară |
8.2. Mușchiul striat scheletic
Mușchiul striat este acel tip de mușchi la care se face referire într-un context general folosind termenul de „mușchi”. O caracteristică de bază a sa este dată de capacitatea lui de a acționa cu o forță asupra oaselor. Contracția celulelor musculare care apare, în general, doar ca urmare a unui stimul (impuls nervos), este un proces activ, iar relaxarea un proces pasiv. Prin contracția mușchilor atașați de anumite părți ale scheletului, contribuie astfel la mișcarea părților respective.
Pentru ca locomoția să aibă loc, este necesar ca două grupuri de mușchi să acționeze unul împotriva celuilalt (mușchi antagoniști), determinând mișcări ale unor părți ale corpului în direcții opuse. Astfel, la articulația genunchiului, flexia posterioară a gambei se realizează de către mușchii flexori, iar extensia anterioară de către mușchii extensori.
Structura țesutului muscular striat scheletic
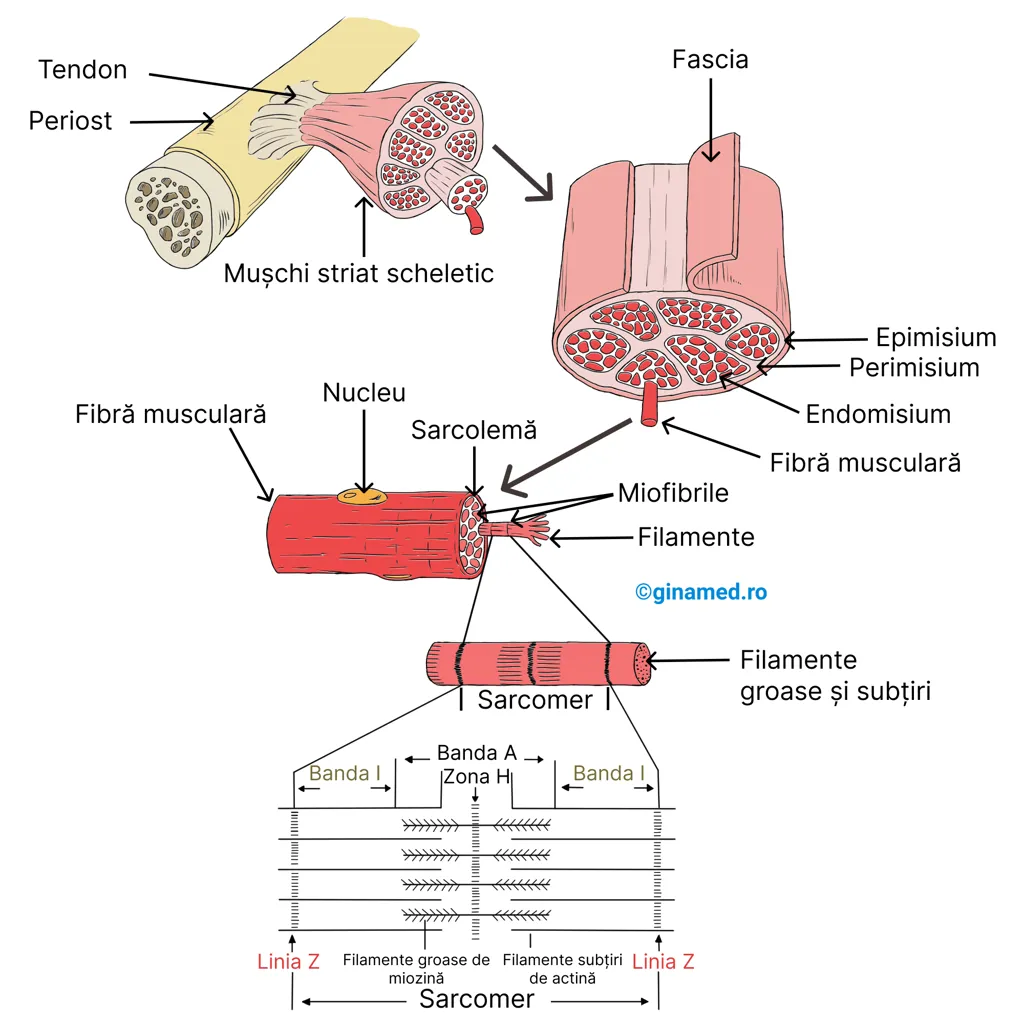
Straturi de țesut conjunctiv învelesc și separă celulele țesutului muscular scheletic astfel:
- endomisium – învelește fiecare fibră musculară;
- perimisium – învelește un pachet de fibre (sau fascicule) musculare; partea din mușchi care conține fibrele musculare este denumită gaster sau corp;
- epimisium și fascia – învelesc mușchiul în întregime.
Fascia superficială este stratul extern al fasciei care la persoanele obeze prezintă o cantitate mare de țesut adipos.
Dincolo de corpul mușchiului, endomisiumul, perimisiumul, epimisiumul și fascia se continuă și formează tendonul care permite atașarea mușchiului de os.
Structura celulei musculare
În alcătuirea mușchiului striat scheletic intră fibre (celule) musculare. În alcătuirea fiecărei fibre musculare intră un set de 4-20 de filamente filiforme numite miofibrile. O miofibrilă are o lățime de 1-2 μ și poate atinge o lungime de 100 μ. Miofibrilele se află în citoplasma celulei musculare denumită sarcoplasmă, în care se mai găsesc și mitocondrii (constituie o sursă de energie sub formă de ATP necesară în contracția miofibrilelor).
De-a lungul axului longitudinal, miofibrilele sunt organizate în unități mai mici cu o lungime de circa 2 μ – sarcomere. Deoarece sunt distribuite în mod repetat, sarcomerele sunt cele care dau un aspect striat caracteristic mușchiului scheletic. Unitatea funcțională a mușchiului striat scheletic este sarcomerul.
Privit la microscop, la nivelul sarcomerului se observă două tipuri de miofilamente distribuite paralel: filamente subțiri (formate din proteina actină) și filamente groase (formate din proteina miozină).
Zone și linii din alcătuirea sarcomerului:
- linia Z – zona în care filamentele de actină din două sarcomere adiacente (apropiate) se întrepătrund; două linii Z delimitează un sarcomer;
- banda I – este o bandă largă, clară, împărțită în două jumătăți egale de către linia Z;
- banda A – este o bandă largă, densă, aflată la mijlocul sarcomerului; rezultă din suprapunerea filamentelor de miozină, printre care se află filamente de actină;
- zona H – este alcătuită doar
din filamente de miozină (actina lipsește); împarte banda A în două
jumătăți egale.
Prin repetiția benzilor A și I se conferă aspectul striat din miofibrilele mușchiului striat.
Funcția mușchilor striați – mecanismul de glisare al filamentelor
Filamentele subțiri de actină sunt ancorate de linia Z. În cursul contracției musculare, filamentele opuse de actină sunt trase de-a lungul filamentelor de miozină. Astfel, sarcomerul se scurtează (distanța dintre liniile Z scade) odată cu creșterea gradului de suprapunere a filamentelor de miozină cu cele de actină. În mușchii scheletici contractați, sarcomerele se scurtează pe măsură ce crește gradul de suprapunere a filamentelor de miozină cu cele de actină.
Molecula proteică de miozină se compune din două lanțuri polipeptidice în care fiecare prezintă o formă asemănătoare unei crose de golf, având axele răsucite una în jurul celeilalte și cu capetele înclinate în lateral ca la nivelul unei balamale. În jurul fiecărui filament de miozină se află filamente subțiri de actină, astfel încât cele două capete ale miozinei și filamentele de actină să vină în contact între ele în cursul contracției.
În mecanismul de glisare al filamentelor, capetele miozinei funcționează ca punți între filamentele de actină și miozină. Capetele aplică un impuls puternic care se aseamănă cu impulsul aplicat de vâsle să împingă apa. Impulsul aplicat trage filamentele subțiri de actină către interior (către zona H) și duce la scurtarea sarcomerului. Atunci când acest proces are loc în același timp în milioane de sarcomere de la nivelul a mii de fibre musculare, are loc scurtarea mușchiului și apariția contracției. Astfel, în starea de contracție musculară filamentele de actină sunt suprapuse, sarcomerele ating dimensiunea lor minimă, zona H dispare și banda I își reduce foarte mult dimensiunile.
Punțile de miozină sunt cele care determină contracțiile fibrelor musculare deoarece ele au un comportament similar unor enzime. Astfel, ATP-ul se fixează de receptorul enzimatic de la nivelul capului miozinei și apoi aceasta este desfăcută (descompusă) în ADP și o grupare fosfat anorganică, care ambele rămân legate de capătul miozinei. Prin descompunerea ATP-ului se eliberează energie care activează capul miozinei în poziție armată, care va determina crearea unei legături slabe între capul miozinic și filamentul de actină. În acest fel se justifică eliberarea ADP-ului și a grupării fosfat, iar capul miozinei stabilește o legătură puternică cu filamentul de actină. Totodată, capul miozinei înaintează și asigură impulsul filamentelor de actină care va determina alunecarea (deplasarea) filamentelor subțiri de actină în lungul celor groase de miozină. Odată cu legarea unei noi molecule de ATP la nivelul receptorului miozinic are loc eliberarea actinei și ciclul discutat anterior se reia atât timp cât persistă stimulul neuronal. În absența unei stimulări nervoase, sarcomerele se relaxează.
Producerea ciclului de glisare a filamentelor se face rapid și are loc la nivelul a milioane de capete miozinice, la fiecare capăt al sarcomerului, în funcție de disponibilitatea ATP-ului care va furniza energie pentru contracție.

Denumirea de fibre musculare roșii este dată de existența a numeroase molecule de mioglobină de culoare roșiatică, la nivelul cărora este stocat oxigenul. Prin urmare, mușchiul roșu mai poartă denumirea de mușchi lent (mușchi oxidativ). În perioada de relaxare sau de contracție lentă a mușchiului, utilizarea ATP-ului este lentă, iar celulele pot genera rapid ATP din oxigenul stocat în mioglobină pentru respirație celulară. Ca urmare a acestor proprietăți, mușchii roșii se pot contracta repetat și totodată pot rezista oboselii musculare.
Un alt tip de mușchi este mușchiul alb, care prezintă o cantitate redusă de mioglobină sau chiar absentă, ceea ce duce la reducerea depozitului de oxigen. Prin abundența conținutului său în glicogen, mușchiului alb i se mai atribuie și denumirea de mușchi rapid (mușchi glicolitic). În acest tip de mușchi, utilizarea ATP-ului este rapidă, însă fără o înlocuire la fel de rapidă, ca urmare a depozitelor scăzute de oxigen disponibile pentru respirația celulară. În acest fel, în mușchiul alb oboseala musculară se instalează rapid cu acumularea de acid lactic.
Inițierea contracției musculare
Filamentele subțiri de actină sunt sub formă de două lanțuri răsucite într-un helix, în șanțul căruia de află molecule de tropomiozină (o proteină). În perioada de relaxare musculară, tropomiozina nu permite legarea dintre capetele de miozină și actină, prin mascarea locului în care această legătura ar fi apărut.
În lungul filamentelor subțiri de actină, în mod regulat, la anumite intervale se întâlnește troponina (o altă proteină). Aceasta se leagă de molecule de tropomiozină, actină și ioni de calciu.
În momentul în care la fibrele musculare ajunge impulsul nervos, acestea se contractă. Procesul este marcat de eliberarea neurotransmițătorului acetilcolină la nivelul joncțiunii neuromusculare (care este alcătuită din: terminația unei singure celule nervoase și o singură fibră musculară). Este important de reținut că cele două membrane ale elementelor care compun joncțiunea neuromusculară sunt foarte apropiate, însă fără să se atingă, lasă un spațiu plin cu lichid denumit fantă sinaptică. În această zonă se eliberează neurotransmițătorii (acetilcolina). Inițierea impulsului are loc în celula musculară după care urmează propagarea sa pe întreaga suprafață celulară. În acest fel, impulsul determină variate evenimente în interiorul celulei musculare care vor duce la contracția ei.
Neurotransmițătorul acetilcolină acționează asupra celulei musculare prin stimularea ei să elibereze ioni de calciu în sarcoplasmă, determinând cuplarea miozinei cu actina și declanșarea contracției.
În totalitatea celulelor în perioada de repaus, transportul activ menține o concentrație scăzută a ionilor de sodiu (se referă la activitatea pompei de sodiu-potasiu). În momentul în care acetilcolina se fixează de receptorii de pe membrana celulară musculară (sarcolemă), se declanșează influxul de ioni de sodiu în celulă.
Totodată, în mod normal, în perioada de repaus celular, există o concentrație foarte scăzută de ioni de calciu în citoplasma celulelor. Acest lucru este menținut prin expulzarea (pomparea) spre exteriorul celulei a ionilor de calciu sau dirijarea lor în interiorul reticulului endoplasmic neted specializat al celulei (denumit și reticul sarcoplasmic). La interiorul fibrelor musculare se află un alt tip de rezervor de calciu care poartă numele de sistemul tubilor transversali (sau tubii T). Tubii T sunt alcătuiți dintr-un sistem de invaginări ale sarcolemei, dispuși la nivelul joncțiunilor A-I și au rolul de a facilita pătrunderea calciului extracelular.
Astfel, prin influxul ionilor de sodiu se produce o activitate electrică în lungul fibrei musculare care va determina tubii T și reticulul sarcoplasmic să elibereze rapid ionii de calciu prin difuziune, din depozite în interiorul sarcoplasmei. În acest fel, ionii de calciu scaldă filamentele de miozină și se fixează la situsurile (locurile) din moleculele de troponină și duce la schimbarea formei complexului. Având în vedere că troponina este legată de tropomiozină, aceasta din urmă își modifică poziția, eliberând locurile de legare (cuplare) ale actinei cu capetele miozinei. În acest fel se declanșează un impuls puternic. În cazul în care se eliberează o cantitate mai mare de ioni de calciu, va rezulta un număr mai mare de legături stabilite cu troponina, permițând în acest fel efectuarea unui număr mai mare de contracții și mai puternice.

Relaxarea musculară
În absența impulsurilor nervoase, mușchiul se relaxează. Astfel, prin stoparea impulsurilor, au loc următoarele fenomene:
- sarcolema și tubii T revin la starea de repaus;
- reticulul sarcoplasmic oprește eliberarea de ioni de calciu;
- enzimele pompează înapoi calciul în cisternele terminale (completare: acestea sunt părți ale reticulului sarcoplasmatic).
- (Completare: Prin "enzimele" de mai sus se face referire la ATP-aza de calciu, pompa care intervine în transportul activ al ionilor de calciu. Este denumită enzimă deoarece catalizează hidroliza ATP-ului și folosește energia din acest proces.)
Prin retragerea calciului, troponina își recapătă configurația de repaus și tropomiozina se distribuie din nou la suprafața locurilor de cuplare ale miozinei de pe filamentele de actină. Aceste filamente alunecă (glisează) către exterior pentru a reda sarcomerului lungimea sa inițială de repaus.
Consum de ATP se face în timpul contracției, dar și relaxării. În perioada de relaxare, ATP-ul oferă energia necesară pompării ionilor de calciu în tubii T și în reticulul sarcoplasmic. Acei mușchi care nu își pot asigura ATP, vor rămâne în stare de contracție. Acest aspect, justifică starea de contracție a celulelor musculare după moarte, denumită rigor mortis. Proteinele contractile (actina și miozina) rămân blocate împreună în poziția de contracție. La interval de câteva ore, ca urmare a altor procese degenerative dominante, mușchii se relaxează.
În timpul relaxării și contracției, în fibra musculară au loc mai multe evenimente surprinse în tabelul de mai jos:
| Etape | Contracția fibrei musculare | Relaxarea fibrei musculare |
|---|---|---|
| 1 | stimulul: eliberarea acetilcolinei de către un neuron motor | membrana musculară nu primește stimuli deoarece colinesteraza descompune acetilcolina |
| 2 | acetilcolina traversează fanta sinaptică | prin transport activ, ionii de calciu sunt aduși în interiorul tubilor T și ai reticulului sarcoplasmic |
| 3 | membrana fibrei musculare este stimulată și impulsul se transmite pe calea tubilor T, în profunzimea fibrei musculare | se rup punțile de legătură dintre filamentele de actină și miozină |
| 4 | are loc difuziunea ionilor de calciu din reticulul sarcoplasmic în sarcoplasmă și apoi se fixează la nivelul moleculelor de troponină | are loc alunecarea (glisarea) filamentelor de actină și miozină, una față de cealaltă |
| 5 | moleculele de tropomiozină se deplasează și etalează (expun) locurile de legare specifice de la nivelul filamentelor de actină | odată cu relaxarea fibrei musculare, aceasta își restabilește lungimea |
| 6 | se stabilesc punți de legătură între filamentele de actină și miozină | interacțiunea dintre filamentele de actină și miozină este blocată de moleculele de troponină și tropomiozină |
| 7 | filamentele de actină glisează spre interior, în lungul filamentelor de miozină | |
| 8 | pe măsură ce contracția are loc, fibra musculară se scurtează |
Răspunsul gradual
O fibră musculară individuală prezintă o caracteristică importantă și anume, un răspuns de tip „tot sau nimic”(adică se contractă complet sau deloc). Cu alte cuvinte, contracția unei fibre musculare se produce doar dacă impulsul depășește o anumită intensitate prag și apoi se răspândește la nivelul întregii celule. În cazul în care valoarea intensității prag a fost depășită sau dacă se continuă creșterea ei, a ratei și a duratei impulsului, contracția produsă nu va fi cu mult mai puternică.
Ca întreg, mușchiul răspunde gradual la stimuli și nu se supune legii „tot sau nimic”. Răspunsul gradual are un caracter variabil, care depinde de numărul de fibre musculare care se contractă într-un mușchi. În situația în care impulsul nervos este condus către mușchi pe un număr crescut de neuroni, atunci în contracție vor fi implicate și mai multe fibre musculare. În caz contrar, prin stimularea unui număr redus de fibre musculare, contracția musculară va fi slăbită. Spre fibrele musculare, fiecare neuron se ramifică, astfel încât un singur neuron poate stimula până la 100 de fibre musculare. Deci, un neuron motor poate deservi un număr variabil de fibre musculare.
O unitate motorie se compune dintr-un neuron (cu funcție motorie – neuron motor) și fibrele (celulele) musculare pe care le stimulează. Prin urmare, un mușchi are mai multe unități motorii.

Secusa este contracția unei singure fibre musculare. Prin stimulare continuă, cresc numărul de secuse, proces denumit sumație.
Sumația prezintă următoarele caracteristici:
- constituie starea în care impulsurile nervoase ajung la un mușchi înainte ca precedenta contracție să fi încetat;
- rezultă parțial din incapacitatea reticulului sarcoplasmic de a recupera ionii de calciu înainte de o nouă stimulare;
- prin sumație, intensitatea unei contracții o depășește întotdeauna pe cea a secuselor individuale; secusele sunt de fapt răspunsuri la stimulări mai rare;
- poate culmina prin tetanus (stare de contracție musculară maximală, susținută); o stare de tetanus muscular se instalează prin strângerea pumnului.
Tonusul muscular (caracterizează mușchii) constituie stare în care mușchiul este menținut parțial în stare de contracție pentru o perioadă mai lungă de timp. Starea de tonus muscular este caracterizată de producerea unei stimulări consecutive a fibrelor musculare, de scurtă durată. În acest fel, anumite zone din mușchi sunt mereu în stare de contracție, comparativ cu cea mai mare parte a sa, care se găsește relaxată.
Sub influența forței gravitaționale, realizarea ortostatismului se face prin intervenția tonusului muscular. Prin exercițiu fizic, tonusul mușchilor implicați este crescut.
8.3. Energia necesară contracției musculare
În vederea realizării contracției musculare, energia folosită (care induce interacțiunea dintre filamentele de actină și miozină) este obținută din ATP. Acesta la rândul său se obține ca urmare a unor reacții chimice care se desfășoară în multitudinea de mitocondrii dispuse în apropierea filamentelor subțiri și groase. ATP-aza, enzima prezentă la nivelul capetelor filamentelor miozinice, scindează ATP-ul în ADP și grupare fosfat, cu eliberare de energie, determinând activitate actino-miozinică.
În fibra musculară există rezerve limitate de ATP, ceea ce necesită o permanentă regenerare de ATP din ADP și grupări fosfat. Fosfocreatina (creatin fosfatul) constituie o sursă de regenerare a ATP-ului. Prin prezența legăturilor sale fosfat cu nivel energetic ridicat, fosfocreatina funcționează ca un depozit de energie celulară. Prin epuizarea ATP-ului, fosfocreatina intervine și eliberează energie, transferând gruparea fosfat unei molecule de ADP și astfel regenerează molecule de ATP.
Epuizarea rezervelor de ATP și fosfocreatină poate avea loc atunci când vorbim de un mușchi extrem de activ, moment în care sursa de energie pentru celula musculară devine metabolismul glucidic. Acesta, prin respirație celulară, implică următoarele procese: glicoliză, ciclu Krebs, sistem transportor de electroni și chemiosmoză. Realizarea reacțiilor respirației celulare necesită oxigen pentru a furniza ATP, reacții care au loc în citoplasmă și mitocondrii.
Distribuirea oxigenului necesar respirației celulare la nivelul fibrelor musculare se face pe calea hemoglobinei eritrocitare. În fibrele musculare, oxigenul se leagă de mioglobină (pigment), care îl depozitează temporar. În acest fel, se reduce necesitatea unui aport continuu de oxigen în timpul contracției, ca urmare a prezenței mioglobinei la nivel muscular.
În cazul unul efort muscular intens timp de câteva minute, este imposibil de asigurat un aport de oxigen suficient de rapid încât să satisfacă necesitățile celulare. Astfel că celulele musculare depind de ATP-ul care rezultă din faza anaerobă a respirației celulare.
În cursul glicolizei anaerobe, moleculele de glucoză trec prin reacții succesive și se convertesc în acid piruvic. Pentru fiecare moleculă de glucoză supusă glicolizei anaerobe, se obțin două molecule de ATP. În cazul în care celula este lipsită de rezerve de oxigen, acidul piruvic este convertit mai departe în acid lactic. Odată cu acumularea acestuia, apare oboseala musculară extremă și datoria de oxigen care presupune suplimentarea nevoilor de oxigen pentru a preveni formarea de acid lactic. Efectul datoriei de oxigen se observă în dificultatea de a respira după un efort intens (extenuant).
Prin prezența sa, acidul lactic modifică pH-ul și astfel fibrele musculare vor avea un răspuns mai slab sub influența stimulilor. Mare parte din acidul lactic produs la nivelul celulelor musculare, difuzează în exteriorul celulei, după care este transportat pe cale sangvină la ficat. Celulele acestuia folosesc oxigenul pentru a reconverti acidul lactic în molecule cu randament energetic ridicat. Totodată, oxigenul este folosit pentru a resintetiza ATP-ul prin respirație celulară și pentru formare de fosfocreatină.

8.4. Funcțiile mușchiului neted și cardiac
Mușchiul neted
Celulele (fibrele) țesutului muscular neted sunt unite între ele prin fibre de colagen și uneori prin joncțiuni de tip „gap”. În citoplasma celulei musculare netede se află numeroase filamente de actină ale căror capete sunt inserate pe suprafața internă a membranei plasmatice. Miozina este și ea prezentă, având capetele miozinice similare celor din mușchiul scheletic. Contracția fibrei musculare netede este mai lentă, comparativ cu fibra musculară scheletică, însă poate persista pentru o perioadă mai lungă de timp (contracție susținută). Mușchiul neted este controlat involuntar, ceea ce îi permite reacții proprii, care se adaptează variatelor condiții, într-un mod inconștient.
În celulele musculare netede, modalitatea de glisare a filamentelor este similară, însă reticulul sarcoplasmic intracelular lipsește, iar calciul are funcție diferită.
Unele fibre musculare netede se află sub coordonarea sistemului nervos vegetativ (autonom), care prin componenta sa simpatică stimulează anumite contracții, pe când componenta sa parasimpatică inhibă anumite contracții. Totodată, fibrele musculare netede sunt reactive și la anumiți hormoni.
Sarcomerele lipsesc din fibra musculară netedă, însă filamente de actină și miozină sunt prezente și organizate în structuri asemănătoare miofibrilelor. Raportul în care se găsesc filamentele de miozină și cele de actină în fibra musculară netedă este de 1:16, comparativ cu 1:2 din fibra musculară scheletică. În plus, fibrele musculare netede conțin filamente intermediare, necontractile care sunt atașate unor structuri denumite corpi denși, distribuiți în întreaga celulă. Corpii denși sunt un echivalent al liniei Z din mușchii scheletici și facilitează cuplarea filamentelor de actină. Corpilor denși le este atașat un citoschelet puternic, alcătuit din filamente intermediare. Țesutul conjunctiv înconjoară fibrele musculare, iar unirea dintre ele se face prin fibre de elastină și colagen. Fibrele musculare netede sunt organizate în straturi musculare (de exemplu, straturile din pereții vaselor sangvine). Prin contracția mușchilor netezi din vezica urinară, rect și uter, aceste organe își pot elibera conținutul. În mușchii netezi, joncțiunile neuromusculare nu sunt bine structurate, însă butonii terminali ai axonilor eliberează neurotransmițători în aceste joncțiuni difuze.
Troponina lipsește din mușchii netezi, deci nu se mai cuplează cu tropomiozina care ar acoperi locurile de cuplare ale actinei cu capetele miozinice și astfel se justifică pregătirea în orice moment a mușchilor netezi pentru contracție. În cazul în care o fibră musculară netedă este activată, intervine calmodulina (nu troponina) și leagă calciul. O serie de reacții duc la fosforilarea capetelor miozinei și permite legarea actinei cu declanșarea contracției. Sistemul nervos vegetativ este cel care trimite impulsuri către mușchii netezi.
Mușchiul neted se clasifică în:
- mușchi neted unitar – fibrele sale se contractă ritmic, ca o unitate; fibrele sunt unite prin joncțiuni de tip „gap";
- mușchi neted multiunitar – fibrele acționează independent una față de cealaltă, sunt rare joncțiuni de tip „gap”; prezintă terminații nervoase bogate, unitatea motorie se formează cu un anumit număr de fibre musculare; mușchiul neted multiunitar se întâlnește în: căile respiratorii, arterele mari, mușchii erectori ai firelor de păr.
Mușchiul cardiac
Doar în structura inimii se întâlnește mușchiul cardiac. Fibrele musculare striate au în alcătuirea lor numeroase mitocondrii, sunt de cele mai multe ori ramificate și formează o adevărată rețea cu fibrele învecinate. Au sistem tubular T, reticul sarcoplasmic și în mod particular, doar în acest tip de țesut muscular sunt prezente discurile intercalare. Acestea sunt regiuni ale membranei celulare care leagă strâns unele de celelalte capetele celulelor cardiace și la nivelul cărora se remarcă numeroase joncțiuni de tip „gap”. Pe calea acestor joncțiuni, impulsul electric se transmite cu ușurință de la o celulă la alta și determină ulterior contracția.
Asupra mușchiului cardiac se exercită un control de tip involuntar, prin impulsuri primite prin ramuri ale sistemului nervos vegetativ care poate modifica contracțiile.
În tabelul de mai jos se compară mușchiul cardiac și cel scheletic:
| Caracteristica | Mușchiul cardiac | Mușchiul scheletic |
|---|---|---|
| Tip de control | sistemul nervos vegetativ | sistemul nervos somatic |
| Dispunerea fibrelor | ramificate | neramificate |
| Aspect microscopic | striat | striat |
| Număr de nuclei/fibră | unul | mulți |
| Discuri intercalare | prezente | absente |
| Aranjamentul tubilor | un tub/sarcomer dispus la nivelul liniei Z | 2 tubi/sarcomer aflați la nivelul joncțiunilor A-I |
| Durata potențialului de acțiune | 150-300 msec | 1-2 msec |
| Timpul de contracție | 150-300 msec | aprox. 40 msec; cu variații la mușchi diferiți |
| Perioada refractară absolută | 150-300 msec | 1-2 msec |
Bibliografie:
- Anatomie și fiziologie umană pentru admiterea la facultățile de medicină; Autori: Krumhardt B., Alcamo I.E.; Barron's & Universitatea de Medicină și Farmacie Târgu Mureș, 2022
